
The brain uses a lot of energy. Neurons communicate with each other electrically via action potentials – spikes – a process which requires a ton of ATP to activate ion pumps on the cell membrane. What the brain uses in energy it must gather it from the outside world; and the world is a scary place, filled with dangers.
The brain is thus under intense evolutionary pressure to use energy efficiently. To do this well requires a tight coupling between action and perception. When a certain part of the visual field is relevant to action, neurons that are sensitive to this part of the visual field respond more robustly. They increase in sensitivity and firing rate – as well as decorrelate – and that generally leads to better performance, in a process called attention.
Attention enhances the representation of a certain – relevant – part of the visual field at the expense of the rest of the visual field. Spikes are better allocated; they’re deployed when they’re needed. I’m sure you’ve all seen attention in action, by now, in this lovely video:
Higher spatial resolution during locomotion
During my postdoc with Dario Ringach at UCLA, we wanted to understand how this process works, so we looked at a related phenomenon in mice; the paper just got published in J Neurosci [PDF]. When a mouse moves around, it exposes itself to danger. There’s a large increase in the gain of neurons in the primary visual cortex of mice during locomotion. It’s as though the visual cortex powers down when the mouse is stationary, and when it’s moving and in danger, the visual cortex turns on.

Under fairly general assumptions about the neural code, increasing gain should mean that stimuli are better represented. We indeed found that all stimuli were better represented during locomotion. This was very tightly coupled with the onset of locomotion (above). The mouse starts moving; visual cortex powers up; and it sees better.

We found, surprisingly, that the increase in gain is concentrated at higher spatial frequencies. What that means is that not only is contrast better represented – blacks are blacker, whites whiter – but the effective spatial resolution of the representation becomes higher. In other words, the mouse sees clearer – and by a wide margin, too.
That’s pretty surprising, and it might help explain a similar phenomenon in human attention – that attention appears to increase spatial resolution at the attended location. In times of need, the brain allocates extra energy to relevant stimuli, and that means that it can perceive visual stimuli more clearly. It’s an efficient and adaptive code.
Optimal decoding is easy
There’s a twist, and it’s a rather subtle, technical one that is buried in our paper. One worry about adaptive coding is that it changes what the neural code means. An adaptive code means that if I measure 2 spikes from a neuron when a mouse is now moving, it means a different thing than when it is moving.
Now if I change the way that I encode stimuli in primary visual cortex, does that mean that every stage higher up must be aware of this? Do I need a new decoder? That would undermine the efficiency of the new code; sure, spikes are more efficiently allocated, but now you twice the neurons downstream need to upkept to read the two different codes.
It turns out that for a very special type of modulation – changing the gain of neurons – you don’t need to do anything special to read the modulated code; you only need one, simple decoder. It falls out of an analysis of optimal decoding of Poisson codes that was pioneered by Jazayeri and Movshon (2006).

Let’s say that I choose to present a stimulus 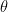

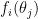
By Bayes’ theorem, the log likelihood of the stimulus, given the observations, is given by:
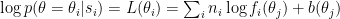
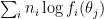
You can interpret the 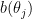
When the neural code is modulated multiplicatively, tuning curves themselves are modified multiplicatively. But what’s important for the decoder is the log of the tuning curves. The multiplicative factor falls out of the 
So we simply need to change the baseline of the decoder to recalibrate it depending on the state of the system. What that means, in this case, is that we have to slightly downvote high spatial frequency signals during locomotion.. That’s very straightforward to do in neural hardware, via, e.g. an efference copy of the locomotion signal and two synapses with different biases in the decoding neural population.
Indeed, empirically, you get as good decoding accuracy with a single decoder that changes its bias during running than with two separate decoders tuned to each condition:

What that implies is that by more efficiently allocating spikes, you get higher spatial resolution when it really matters, without needing special hardware to understand the adaptive neural code.
The paper is out in J Neurosci now:
P. J. Mineault, E. Tring, J. T. Trachtenberg, D. L. Ringach (2016). Enhanced Spatial Resolution During Locomotion and Heightened Attention in Mouse Primary Visual Cortex. J Neurosci. 36(24):6382– 6392. [PDF]

One response to “Enhanced spatial resolution during locomotion”
Nice! Reminds me some of the result in the microsaccade literature such as http://www.sciencedirect.com/science/article/pii/S0960982212000802
A small comment: did you try to perform a log transform on the x-axis for figure 3B? It might arguably be better for spatial frequency data.