
Adam delivered a pretty intense lecture at CSHL on population coding, correlations and phase-locking. Consider myself mindfucked. Mainen & Sejnowski (1995) showed that single neurons have very reliable responses to current injections. Nevertheless, cortical neurons seem to have Poisson or supra-Poisson variability. It’s possible to find a bound on decodability using the Fisher information matrix (Sompolinsky & Seung 1993). Under the assumption of independent Poisson variability, it is possible to derive a simple scheme for ML decoding that can be implemented in neuronal populations (Jazayeri & Movshon 2006). Liam’s class notes on decoding are relevant here.
In reality, the accumulation of noise sources and various other (poorly understood) mechanisms cause cortical neuronal populations to be correlated, which poses challenges for decoding. You can get a little more juice out of decoding algorithms by considering pairwise correlations (Pillow et al. 2008). Schneidman et al. (2006) argue that correlations are due to pairwise, but not 3-wise interactions between neurons, although Adam hinted that closer inspection reveals otherwise.
Correlations between neurons can occur at various timescales. It’s possible, by integrating the cross-correlation function (xcorr in Matlab) between two neurons, to read off the timescale of the correlation (Bair, Zohary and Newsome 2001). This can help to distinguish correlations due to monosynaptic or disynaptic connections, which are visible at short timescales, with correlations due to slow drift in oscillations, up-down states, attention, etc., which occur at much longer timescales. Correlations depend on physical distance on the cortical map as well as tuning distance between two neurons (Smith and Kohn, 2008).
Gamma oscillations are one cause of correlated firing; firing is more likely during certain phases of the gamma oscillation. If an output population has an intrinsic phase 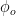

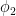
This sounds handwavy to me, and indeed Adam showed data that is inconsistent with this hypothesis. A Utah array was implanted in V1 while a linear array was used in V2 during visual stimulation and spontaneous activity.
Neurons tend to be phase locked to the gamma rhythm. It’s known that the strength of gamma decreases and low frequency increases with stimulus strength. Because the larger part of spike-spike correlations are due to low frequency phase locking, spike-spike correlations should decrease with stimulus strength, and indeed this is the case, although it’s unclear whether the numbers add up, so to speak.
The question then is whether fluctuations in excitability due to the V2 gamma rhythm have any effect on V1-V2 coupling. This can be done by looking at the probability of a spike in V2 appearing a short time window after a V1 spike conditional on the LFP phase in either V1 or V2. It appears that the efficiency of the transmission is modulated by the V1 phase but not the V2 phase, which is inconsisten with the Fries hypothesis.
One interesting observation is that the mean phase of the gamma rhythm is delayed by 90 degrees between V1 and V2. However, the mean latency between V1 and V2, which can be read off of the mean cross-correlation function in V1-V2 pairs, is about 2.4 ms. Assuming a gamma oscillation peaking at 40 Hz, that means that the phase offset, if the gamma rhythm is driven by V1 and amplified by V2, should be about 30-40 degrees.
Thus, it seems as though the gamma rhythm is not amplified but rather suppressed or smeared (so to speak) in V2, and that goes against a positive role for gamma [[explain more]]. Rather, it argues for the notion that gamma is an epiphenomenon created by interactions between excitation and inhibition, something which appears consistent with hippocampal slice experiments (say, Traub et al. 1996).
While I was researching this post, I stumbled upon a paper named How Connectivity, Background Activity, and Synaptic Properties Shape the Cross-Correlation between Spike Trains which looks quite relevant. I had a quick talk with Adam afterward and he mentioned Renart et al. (2010) and Rajan et al. (2010) as relevant to networks with balanced excitation and inhibition.
